Corlett, P.R., Mollick, J.A. & Kober, H.
Neuropsychopharmacol. (2022).
https://doi.org/10.1038/s41386-021-01264-3
Abstract
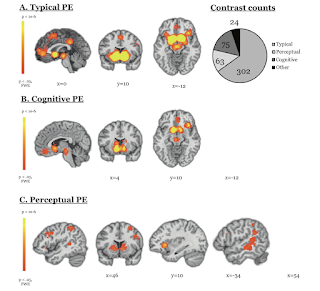
Prediction errors (PEs) are a keystone for computational neuroscience. Their association with midbrain neural firing has been confirmed across species and has inspired the construction of artificial intelligence that can outperform humans. However, there is still much to learn. Here, we leverage the wealth of human PE data acquired in the functional neuroimaging setting in service of a deeper understanding, using an MKDA (multi-level kernel-based density) meta-analysis. Studies were identified with Google Scholar, and we included studies with healthy adult participants that reported activation coordinates corresponding to PEs published between 1999–2018. Across 264 PE studies that have focused on reward, punishment, action, cognition, and perception, consistent with domain-general theoretical models of prediction error we found midbrain PE signals during cognitive and reward learning tasks, and an insula PE signal for perceptual, social, cognitive, and reward prediction errors. There was evidence for domain-specific error signals––in the visual hierarchy during visual perception, and the dorsomedial prefrontal cortex during social inference. We assessed bias following prior neuroimaging meta-analyses and used family-wise error correction for multiple comparisons. This organization of computation by region will be invaluable in building and testing mechanistic models of cognitive function and dysfunction in machines, humans, and other animals. Limitations include small sample sizes and ROI masking in some included studies, which we addressed by weighting each study by sample size, and directly comparing whole brain vs. ROI-based results.
Discussion
There appeared to be regionally compartmentalized PEs for primary and secondary rewards. Primary rewards elicited PEs in the dorsal striatum and amygdala, while secondary reward PEs were in ventral striatum. This is consistent with the representational transition that occurs with learning. We also found separable PEs for valence domains: caudal regions of the caudate-putamen are involved in the learning of safety signals and avoidance learning, more anterior striatum is selective for rewards, while more posterior is selective for losses. We found posterior midbrain aversive PE, consistent with preclinical findings that dopamine neurons––which respond to negative valence––are located more posteriorly in the midbrain and project to medial prefrontal regions. Additionally, we found both appetitive and aversive PEs in the amygdala, consistent with animal studies. The presence of both appetitive and aversive PE signals in the amygdala is consistent with its expanding role regulating learning based on surprise and uncertainty rather than fear per se.
Perhaps conspicuous in its absence, given preclinical work, is the hippocampus, which is often held to be a nexus for reward PE, memory PE, and perceptual PE. This may be because the hippocampus is constantly and commonly engaged throughout task performance. Its PEs may not be resolved by the sluggish BOLD response, which is based on local field potentials and may represent the projections into a region (and therefore the striatal PE signals we observed may be the culmination of the processing in CA1, CA3, and subiculum). Furthermore, we have only recently been able to image subfields of the hippocampus (with higher field strengths and more rapid sequences); as higher resolution PE papers accrue we will revisit the meta-analysis of PEs.